Cell viability is one of the most widely used evaluation metrics in cell-based research; however, the concept is often oversimplified in interpretation. Viability is generally understood as a measure of cell survival, but in practice, most assays and staining methods do not directly measure cell survival.
Instead, these methods generate signals based on specific biological properties such as membrane integrity, nucleic acid accessibility, intracellular enzymatic activity, and metabolic competence. These parameters are independent of one another and change asynchronously during different stages of cell death.
For example, in the early stages of apoptosis, metabolic alterations may occur while the plasma membrane remains intact, whereas in necrosis, loss of membrane integrity is a defining feature. As a result, even within the same cell population, different assays or stains can yield different outcomes depending on the biological parameter being measured.
In this context, cell viability staining should not be considered merely as a tool for distinguishing live and dead cells, but rather as an analytical approach that selectively reflects specific biological endpoints.
Therefore, the selection of staining methods should not be based solely on determining cell survival, but on a clear definition of the biological parameter of interest.
In this article, cell viability stains are categorized based on their underlying mechanisms and measured targets, and their biological significance and applications are discussed.
These conceptual differences can be more intuitively understood through the following figure.
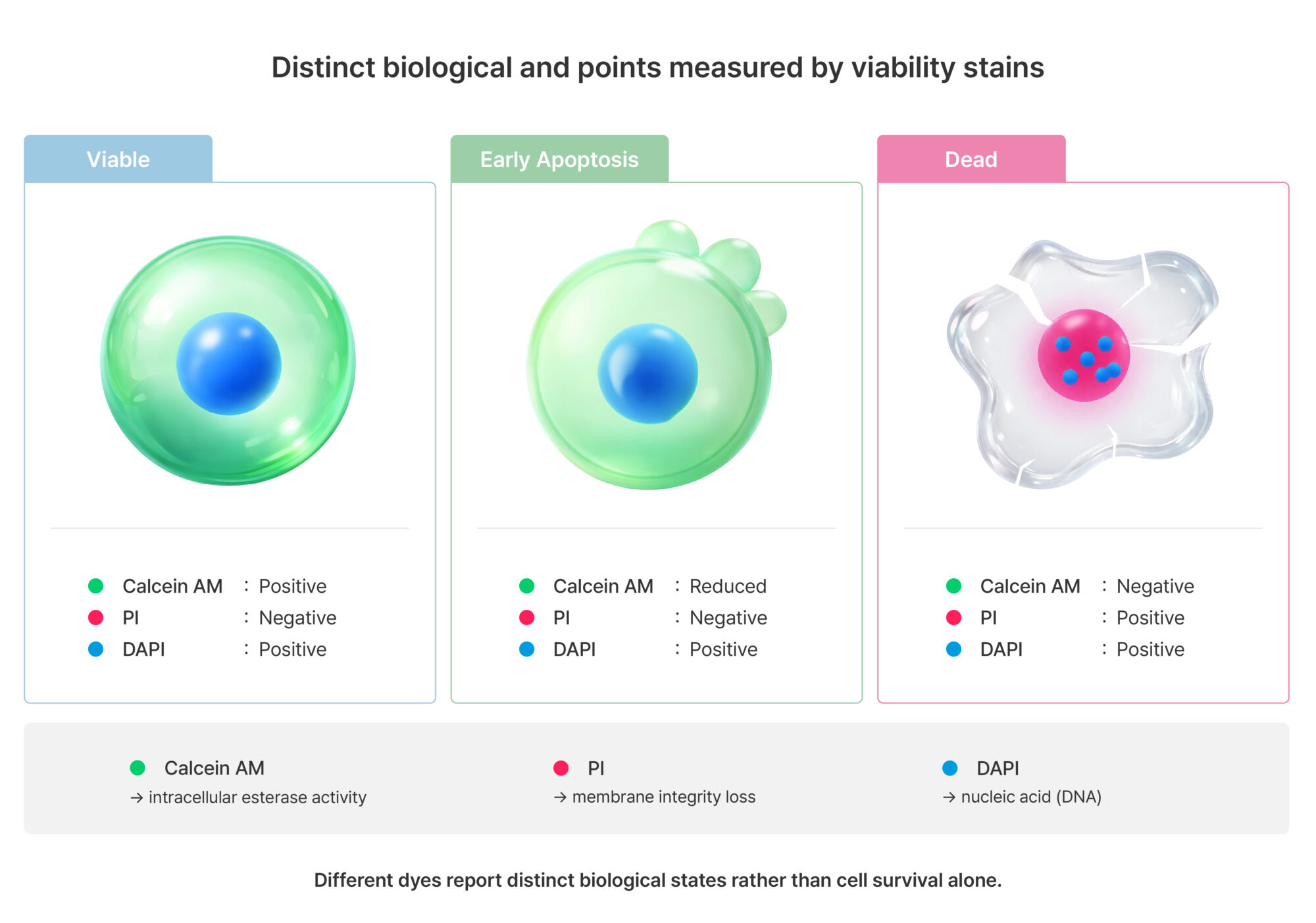
Figure 1. Distinct biological endpoints measured by cell viability stains
Cell viability staining utilizes various dyes that reflect cell states based on different biological parameters. Calcein AM generates fluorescence signals in live cells based on intracellular esterase activity, whereas propidium iodide (PI) enters cells only when membrane integrity is compromised and binds to nucleic acids. DAPI stains the nucleus regardless of cell viability. Due to these differences, even within the same cell population, different dyes can produce different outcomes. Therefore, staining results should not be interpreted as a direct measure of cell survival, but rather as indicators reflecting specific biological states.
Representative dyes include Trypan blue and Erythrosin B.
Trypan blue and Erythrosin B function based on the selective permeability of the cell membrane and are preferentially taken up by cells with compromised membrane integrity. In contrast, viable cells maintain an intact plasma membrane, which restricts dye entry; therefore, live cells appear unstained.
Due to these characteristics, these dyes are widely used as the simplest method for assessing viability based on membrane integrity, particularly in cell counting applications.
However, this approach reflects only the state after membrane damage has occurred and does not capture early apoptotic stages or functional changes such as metabolic alterations. In addition, because it relies on a single parameter, the information obtained about cell state is inherently limited.
Representative dyes include Acridine Orange (AO), DAPI, and SYTO9.
These dyes penetrate the cell membrane and bind to intracellular nucleic acids, staining all cells regardless of their viability. This binding occurs through intercalation or electrostatic interactions, and fluorescence signals can be used to quantitatively analyze nuclear position, morphology, and total cell number.
Due to these characteristics, these dyes are well suited for analyzing entire cell populations and for imaging-based segmentation, and are widely used in high-content analysis.
However, nucleic acid staining does not reflect membrane integrity or the functional state of the cell, and therefore is not suitable as a standalone indicator of viability. As a result, these dyes are typically used in combination with cell-impermeable dyes or dyes that reflect functional parameters.
Representative dyes include Propidium iodide (PI), SYTOX, and Ethidium homodimer (EthD).
These dyes possess physicochemical properties that prevent them from crossing an intact membrane and therefore enter cells only when membrane integrity is compromised. Once inside the cell, they bind to nucleic acids and generate strong fluorescence signals, allowing selective detection of cells with damaged membranes.
Propidium iodide (PI) is a representative DNA intercalating dye and is widely used in fluorescence imaging and flow cytometry. It exhibits high specificity for detecting dead cells and can be applied using relatively simple staining protocols.
However, because PI staining is based on membrane integrity, no signal is observed during the early stages of apoptosis. In addition, its ability to bind RNA can lead to increased background signals if RNase treatment is not performed, and signal variability may occur depending on experimental conditions.
Therefore, PI should not be interpreted as a marker of overall cell death, but rather as an indicator of late-stage apoptosis or necrosis.
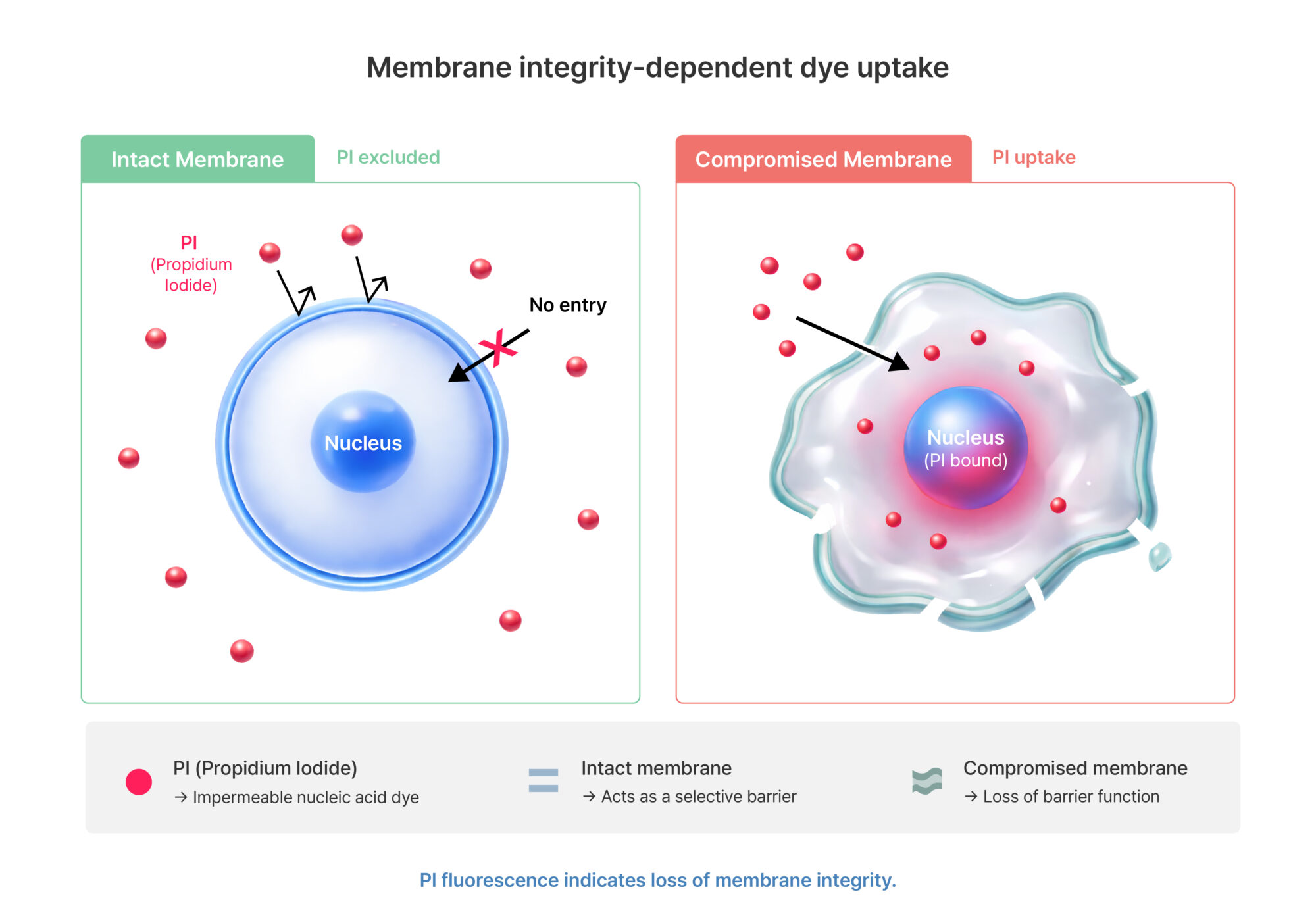
Figure 2. Membrane integrity-dependent dye uptake
Impermeable nucleic acid dyes such as propidium iodide (PI) cannot penetrate an intact plasma membrane and therefore do not enter cells when membrane integrity is preserved. In contrast, when membrane integrity is compromised, these dyes diffuse into the cell, bind to nucleic acids, and generate fluorescence signals.
Due to these characteristics, PI staining should be interpreted not as a direct measure of cell survival, but as an indicator of the loss of membrane integrity.
Representative dyes include Fluorescein diacetate (FDA) and Calcein AM.
These dyes enter cells in a non-fluorescent form and are subsequently hydrolyzed by intracellular esterases into fluorescent products. This process occurs only when both membrane integrity and intracellular enzymatic activity are maintained.
For example, Calcein AM is converted into calcein following esterase-mediated cleavage, and accumulates in the cytoplasm, producing a stable fluorescence signal.
Due to these properties, these dyes enable selective detection of live cells and are useful for assessing functional cellular states.
However, esterase activity is influenced by metabolic conditions, and therefore a decrease in fluorescence signal does not necessarily indicate cell death. For instance, fluorescence intensity may decrease under metabolic suppression even when cells remain viable.
Thus, these dyes should be interpreted as indicators of functional viability or metabolic competence, rather than as direct measures of cell survival.
Each staining method reflects different biological parameters; therefore, it is important to select appropriate dyes or use them in combination depending on the experimental objective.
Analysis based on a single dye may only partially reflect the cellular state and has limitations in distinguishing between different biological processes such as cytotoxicity, cytostasis, and metabolic suppression.
Accordingly, the following combinations are commonly used:
Cell viability staining is not a single analytical concept, but rather a collection of approaches that reflect different biological states. Each dye provides information within a limited scope based on its specific mechanism of action, and even under identical experimental conditions, different staining methods may yield different results.
Therefore, interpreting signals obtained from a specific dye as a direct measure of cell survival may lead to an oversimplification of biological complexity.
For more reliable analysis, the following considerations are essential:
For more information, please visit www.logosbio.com.
References
Looking for the Korean version? Click here.





